III.- Procesamiento de la información visual en la Retina
En este capítulo vamos a tratar las bases neurológicas de la visión a nivel retiniano y para ello veremos el proceso de la fotoquímica y las transducción en la retina, los fenómenos de convergencia de los foto-receptores sobre las células ganglionares y las redes neurales de la inhibición lateral, que permite explicar la percepción de contrastes y que constituye el primer punto clave en la visión de los objetos.
PROCESAMIENTO NEURAL EN LA RETINA.
FOTOQUÍMICA DE LA VISIÓN:
La atmósfera terrestre solo permite ser atravesada por radiaciones entre 300 y 1100 nm y longitudes de onda superiores a 850 nm, sus cuantos de engría, tienen un nivel energético insuficiente para isomerizar las moléculas orgánicas. Por el otro lado, energías inferiores a 300 nm, pueden llegar a destruir algunas proteínas.
El sistema visual utiliza la banda entre 380 y 780 nm. Una reacción fotoquímica consiste en la acción de un fotón (cuanto luminoso), sobre un átomo, excitaría los electrones haciéndolos saltar a una órbita más periférica, sobreelevando la energía del átomo, hasta provocar la escisión de la molécula.
La reacción fotoquímica en la retina consiste en la acción de los fotones de luz sobre los pigmentos de los foto-receptores, provocando la hiperpolarización de sus membranas externas. A 20º de la fóvea (zona de máxima sensibilidad), con una longitud optima de 510 nm, se sabe que la mínima energía que debe incidir sobre el ojo para que se genere el efecto fotoquímico y se inicie una señal visual es de, 2,1 x 10 -10 hasta 5,7 x 10 -10 , según la especie. Considerando la absorción y reflejo de la luz en la córnea y en otros tejidos del ojo, sabemos que bastaran de 6 a 14 fotones para iniciar la señal de activación de los foto-receptores.
KUHNE (1879) fue el primero que aisló una sustancia fotosensible en la retina, localizada en el segmento externo de los bastones, llamándola ERITROPSINA por su color rojo anaranjado brillante. Se le puso RODOPSINA al utilizarse el prefijo griego RODHOS que significa rosado.
La rodopsina es una proteína conjugada incluida en la doble capa lipidia del articulo externo de los bastones, en sus discos. Está compuesta por la glucoproteina OPSINA y el isómero 11- cis del aldehido de la Vit A o RETINAL, con un peso molecular de 27.000 y 41.000 daltons, respectivamente. Cuando se expone a la luz la molécula de rodopsina , el 11cis retinal, cuya estructura tiene forma acodada y está ligada a la opsina, experimenta la transformación a una configuración rectilínea, RETINAL TODO TRANS. La isomerización del retinal va seguida de la disociación de la molécula en opsina libre y todo trans.
Cuando un fotón es absorbido por la rodopsina esta se decolora rápidamente, se activa, el retinal pasa de la forma cis a trans y se escinden las moléculas, provocando la hiperpolarización de la membrana externa del foto-receptor, siguiendo una secuencia bioquímica que se conoce como fototransducción y que se muestra en la figura:

FOTOTRANSDUCCIÓN:
Cuando el fotón llega a la retina debe ser absorbido por el foto-receptor, la señal será ampliada para que sea plenamente efectiva, mediante la cascada enzimática. Las reacciones bioquímicas que tendrán lugar acaban con la hidrólisis del GMP cíclico y el cierre de los canales del sodio y el calcio. La acción del fotón en el foto-receptor desencadena la hiperpolarización de su segmento externo, es lo que denominamos POTENCIAL DE REPOSO, y tiene diferente duración en conos y bastones, pudiendo prolongarse hasta 1 seg, lo cual explicaría que una imagen que se proyecta sobre la retina durante una millonésima de segundo, puede producir la sensación de que sigamos viendo esa imagen durante más de un segundo (postimagen).
En la mayoría de células del organismo, existe una mayor concentración de sodio en el exterior respecto al interior y al contrario en el caso del potasio, más concentrado en el interior respecto al exterior. Este gradiente se mantiene gracias a la acción de la enzima sodio-potasio ATPasa. En el foto-receptor ocurre una situación diferente. En la oscuridad, la membrana plasmática del segmento externo es muy permeable al sodio, mientras que el segmento interno, su membrana plasmática, es muy poco permeable al sodio y mucho más al potasio.
El sodio entra en el segmento externo a través de los canales del sodio (proteínas de membrana), difunde hacia el segmento interno y vuelve a salir hacia fuera por acción de la enzima ATPasa. Se establece lo que llamamos corriente oscura (HAGINS 1970). La entrada de sodio es lo que provoca la despolarización en el foto-receptor, manteniendo abiertos los canales de calcio que existen en el botón sináptico. Esto produce una liberación constante de neurotransmisor, glutamato, hacia la célula bipolar.
Cuando llega luz a la retina se bloquea este flujo de iones. De forma casi exponencial, se frena la entrada de sodio desde el exterior, esto supone que el interior de la membrana se torna más electronegativo. El sodio sale por los segmentos internos y ya no entra por el segmento externo, se produce una hiperpolarización, es decir, disminución de la corriente oscura, así se reduce la liberación de neurotransmisor en la sinapsis (glutamato) y se genera una señal que dará como resultado la génesis de potenciales de acción en las células ganglionares.
La acción de la luz reduce la concentración de GMP cíclico, produciéndose el bloqueo en la entrada del sodio. Asimismo se produce un bloqueo en la entrada de calcio. El aumento en la concentración de sodio en el exterior activa lo que conocemos como BOMBA INTERCAMBIADORA DE SODIO-CALCIO, lo cual provoca la liberación de calcio al exterior desde el segmento externo. La reducción del calcio intracelular inhibe la granulatociclasa y activa la fosfodiesterasa, así se vuelven a abrir los canales para el sodio y calcio, recuperándose el foto-receptor tras la excitación lumínica, dejándolo listo para la llegada de un nuevo fotón.

PROCESAMIENTO NEURONAL MEDIANTE LA CONVERGENCIA.
Al observar las conexiones de conos y bastones con otras neuronas de la retina, nos daremos cuenta que hay un nivel de convergencia diferente en cada uno de estos foto-receptores. La convergencia de los bastones es mayor que la de los conos. Se admite una media de 120 bastones convergiendo sobre una sola célula ganglionar, mientras que tan solo seis conos convergen sobre una ganglionar, esta reducción es incluso mayor en la fóvea, donde llegamos a tener conexiones unitarias, un cono una ganglionar, es decir ya no habría convergencia, sería una conexión directa en la fovea. Esta diferencia entre conos y bastones explica que la visión mediada por conos sea más precisa en la detección de detalles, mientras que los bastones tienen mejor sensibilidad al contraste y menos sensibilidad al detalle, ahora veremos esto con detalle.
Los bastones son más sensibles que los conos al contraste porque necesitan menos luz para generar una respuesta y, especialmente por el fenómeno de convergencia, que determina una sumación de las intensidades. Si tenemos dos células ganglionares que necesitan cada una de ellas 5 unidades de intensidad para activarse, y sobre la primera tenemos un conjunto de cinco bastones convergiendo sobre ella y sobre la segunda le llega la conexión de un solo cono, cuando lleguen estímulos de 1 unidad de intensidad, por ejemplo ocupando un área que activa 5 foto-receptores, los bastones se estimularan y cada uno de ellos enviará 1 unidad sobre la cl ganglionar de convergencia, sumando 5 unidades, el mínimo requerido para activarla, mientras que en el caso de los conos, al activarse, envían tan solo una señal de 1 unidad y, si el estímulo activa 5 conos, como cada uno conecta con una cl. ganglionar, siempre le llega a la cl. ganglionar 1 unidad, no hay efecto de sumación como en los bastones ya que no hay convergencia múltiple, por tanto ese estímulo sí excita la célula ganglionar de los bastones pero no las células ganglionares de los conos. Esto explica porque los bastones son más sensibles a la luz que los conos.
Cuando analizamos la capacidad de detectar detalles, vemos que el fenómeno de convergencia produce el efecto contrario. En los conos de la fóvea, gracias a que cada uno conexiona con una ganglionar, permite que la resolución de la información sea mayor, podemos analizar la imagen punto por punto mientras que los bastones, como convergen varios en una ganglionar, la información se diluye y la resolución baja. Esto determina el concepto de agudeza visual, o capacidad para percibir los detalles.
En la oscuridad, la agudeza visual disminuye porque los conos dejan de funcionar y la visión se debe sólo a la acción de los bastones, con menor capacidad de resolución que los conos, por ello vemos menos nítido.
PROCESAMIENTO NEURAL MEDIANTE EXCITACIÓN E INHIBICIÓN.
Todos los sistemas neuronales funcionan con redes neurales que forman circuitos, es decir, un conjunto de neuronas conectadas entre sí. Cuando estamos en la situación de la fóvea, donde cada cono tiene su ganglionar, la activación de un cono-ganglionar, no se ve afectada por la estimulación de otros conos contiguos, mientras que en la retina periférica, donde predominan los fenómenos de convergencia, la activación de un bastón o más bastones contiguos, sí determina variaciones en la ganglionar donde convergen así, cuantos más bastones se estimulen, mayor frecuencia de respuesta en la ganglionar, tal como se observa en la figura.

Los circuitos pueden complicarse, especialmente cuando aparecen neuronas excitadoras, como el ejemplo anterior, junto a neuronas inhibidoras, como en la figura siguiente, donde las ganglionares A y C bloquean a la ganglionar B. Si el estímulo cae sobre los receptores centrales, 3-4-5, se activa la ganglionar B y se da un efecto excitador pero, si el estímulo es mayor en superficie y se excitan los foto-receptores 2 y 6, se activa parcialmente las ganglionares A y C y producen un efecto inhibidor sobre la ganglionar B que reduce su activación. Si el tamaño del estímulo aumenta y activa a todos los receptores, se suman los 1 y 7 y se activa con mayor potencia las ganglionares A y C, con lo que el efecto inhibidor sobre la B, es más potente y la respuesta total es menor.

Recordemos que la retina ésta formada por diversos tipos de células y, la activación del receptor, cono o bastón, genera estímulos que llegan a las ganglionares a través de las células bipolares, horizontales y amacrinas, lo cual permite pensar que se pueden crear circuitos muy diversos, con vías de excitación o inhibición. Estas posibilidades permiten entender el concepto de campo receptor. En el ejemplo anterior, tendríamos un campo receptor correspondiente a una cel. ganglionar, en este caso la B, del tipo centro ON y periferia OF, también llamada on-of o centro-periferia, ya que si se estimula la zona central, se da una respuesta positiva de activación pero, si se actica la zona periférica, se inhibe la respuesta de la B y, si se activan todos los receptores de esta ganglionar B, se da una respuesta mínima, ya que la excitación de la zona central se ve inhibida por la zona periférica. Este tipo de respuesta, centro periferia, puede darse de la manera que acabamos de ver o, algo más complicada, las ganglionares A y C pueden no contactar directamente con la B, y hacerlo indirectamente mediante células de conexión, como las horizontales y amacrinas, que transmiten el efecto inhibidor, como el ejemplo anterior, pero nos permitirán entender mejor lo que conocemos como inhibición lateral, clave para la detección de contrastes.

La inhibición lateral fue demostrada en 1956 por Hatline, Wagner y Ratliff, utilizando el cangrejo de la especie Limulus y permitió demostrar como la estimulación de receptores adyacentes puede inhibir la respuesta de un receptor central, tal como se muestra en la figura.
PROCESAMIENTO NEURONAL Y PERCEPCIÓN.
El fenómeno de la inhibición lateral permite explicar fenómenos como el de la rejilla de Hermann, donde al mirar la rejilla en conjunto, vemos que en las intersecciones aparecen puntos grises, puntos que desaparecen si nos fijamos directamente en la intersección, demostrando que no son reales.

En la figura vemos que la ganglionar A, que está en medio de los dos pasillos, le llega inhibición de las 4 ganglionares que la rodean, mientras que la ganglionar B, que está en medio de un solo pasillo, le llega inhibición de las ganglionares que están en el pasillo, pero no de las ganglionares que están tapadas por los cuadrados negros, por ello la respuesta de la B es mayor que la de A y esto determina que en A se vea menos brillante y aparece como un punto gris en medio de la intersección de los cuatro cuadrados negros.

Otro de los efectos que se explican por la inhibición lateral son las Bandas de Mach. Tal como se muestra en la figura, si sobre una lámina iluminada colocamos algo que hace sombra, la línea divisoria de la zona en sombra respecto a la zona que sigue iluminada, no la percibimos como una línea bien definida sino que se aprecia una estrecha banda clara y, en la zona clara, junto a la línea divisoria, se ve una banda oscura, sin embargo, el análisis con fotómetro muestra que no hay tales bandas, registra una separación brusca y bien definida entre la zona más oscura y la más clara, lo que nosotros vemos, estas bandas de separación es un fenómeno subjetivo conocido como bandas de Mach y se deben al fenómeno de inhibición lateral.



Las bandas de Mach las podemos explicar con una representación en la que imaginamos 6 receptores, 3 estimulados por la zona clara y 3 por la zona oscura, de forma que cada receptor envía señales de inhibición lateral a los receptores contiguos. Si los receptores de la banda clara, supongamos que generan una respuesta de valor 100 y, los de la banda oscura, de valor 20. Si asumimos que cada célula envía una inhibición de una décima parte del valor de su respuesta, en las ganglionares de la banda clara, tendrán un valor de 10, mientras en las de la banda oscura, de 2. Si calculamos la respuesta final, resta de la activación menos la inhibición, en la ganglionar A, tendrá un valor de 100-10-10=80, en la B, igual, 80, pero en la C, 100-10-2= 88, en la D: 20-10-2= 8 y en E y F, 20-2-2= 16. En las ganglionares del borde, la inhibición es diferente, en la C, de la banda clara, hay menos inhibición, 88, respecto a los 80 de A y B, lo que hace que se vea más brillante y, en el otro lado, la ganglionar D, tiene mayor inhibición que las E y F, ya que recibe la inhibición de C, con valor de 10, lo cual hace que la respuesta final sea de 8, frente a los 16 de las otras, lo cual hace que se vea una franja más oscura.
El fenómeno de la inhibición lateral permite establecer un mecanismo fisiológico para hacer resaltar los bordes y también permite explicar los fenómenos de ilusiones ópticas como el contraste simultaneo de claridad, la cruz de Benary o la ilusión de White.
PROCESAMIENTO NEURAL EN LA RETINA.
FOTOQUÍMICA DE LA VISIÓN:
La atmósfera terrestre solo permite ser atravesada por radiaciones entre 300 y 1100 nm y longitudes de onda superiores a 850 nm, sus cuantos de engría, tienen un nivel energético insuficiente para isomerizar las moléculas orgánicas. Por el otro lado, energías inferiores a 300 nm, pueden llegar a destruir algunas proteínas.
El sistema visual utiliza la banda entre 380 y 780 nm. Una reacción fotoquímica consiste en la acción de un fotón (cuanto luminoso), sobre un átomo, excitaría los electrones haciéndolos saltar a una órbita más periférica, sobreelevando la energía del átomo, hasta provocar la escisión de la molécula.
La reacción fotoquímica en la retina consiste en la acción de los fotones de luz sobre los pigmentos de los foto-receptores, provocando la hiperpolarización de sus membranas externas. A 20º de la fóvea (zona de máxima sensibilidad), con una longitud optima de 510 nm, se sabe que la mínima energía que debe incidir sobre el ojo para que se genere el efecto fotoquímico y se inicie una señal visual es de, 2,1 x 10 -10 hasta 5,7 x 10 -10 , según la especie. Considerando la absorción y reflejo de la luz en la córnea y en otros tejidos del ojo, sabemos que bastaran de 6 a 14 fotones para iniciar la señal de activación de los foto-receptores.
KUHNE (1879) fue el primero que aisló una sustancia fotosensible en la retina, localizada en el segmento externo de los bastones, llamándola ERITROPSINA por su color rojo anaranjado brillante. Se le puso RODOPSINA al utilizarse el prefijo griego RODHOS que significa rosado.
La rodopsina es una proteína conjugada incluida en la doble capa lipidia del articulo externo de los bastones, en sus discos. Está compuesta por la glucoproteina OPSINA y el isómero 11- cis del aldehido de la Vit A o RETINAL, con un peso molecular de 27.000 y 41.000 daltons, respectivamente. Cuando se expone a la luz la molécula de rodopsina , el 11cis retinal, cuya estructura tiene forma acodada y está ligada a la opsina, experimenta la transformación a una configuración rectilínea, RETINAL TODO TRANS. La isomerización del retinal va seguida de la disociación de la molécula en opsina libre y todo trans.
Cuando un fotón es absorbido por la rodopsina esta se decolora rápidamente, se activa, el retinal pasa de la forma cis a trans y se escinden las moléculas, provocando la hiperpolarización de la membrana externa del foto-receptor, siguiendo una secuencia bioquímica que se conoce como fototransducción y que se muestra en la figura:

FOTOTRANSDUCCIÓN:
Cuando el fotón llega a la retina debe ser absorbido por el foto-receptor, la señal será ampliada para que sea plenamente efectiva, mediante la cascada enzimática. Las reacciones bioquímicas que tendrán lugar acaban con la hidrólisis del GMP cíclico y el cierre de los canales del sodio y el calcio. La acción del fotón en el foto-receptor desencadena la hiperpolarización de su segmento externo, es lo que denominamos POTENCIAL DE REPOSO, y tiene diferente duración en conos y bastones, pudiendo prolongarse hasta 1 seg, lo cual explicaría que una imagen que se proyecta sobre la retina durante una millonésima de segundo, puede producir la sensación de que sigamos viendo esa imagen durante más de un segundo (postimagen).
En la mayoría de células del organismo, existe una mayor concentración de sodio en el exterior respecto al interior y al contrario en el caso del potasio, más concentrado en el interior respecto al exterior. Este gradiente se mantiene gracias a la acción de la enzima sodio-potasio ATPasa. En el foto-receptor ocurre una situación diferente. En la oscuridad, la membrana plasmática del segmento externo es muy permeable al sodio, mientras que el segmento interno, su membrana plasmática, es muy poco permeable al sodio y mucho más al potasio.
El sodio entra en el segmento externo a través de los canales del sodio (proteínas de membrana), difunde hacia el segmento interno y vuelve a salir hacia fuera por acción de la enzima ATPasa. Se establece lo que llamamos corriente oscura (HAGINS 1970). La entrada de sodio es lo que provoca la despolarización en el foto-receptor, manteniendo abiertos los canales de calcio que existen en el botón sináptico. Esto produce una liberación constante de neurotransmisor, glutamato, hacia la célula bipolar.
Cuando llega luz a la retina se bloquea este flujo de iones. De forma casi exponencial, se frena la entrada de sodio desde el exterior, esto supone que el interior de la membrana se torna más electronegativo. El sodio sale por los segmentos internos y ya no entra por el segmento externo, se produce una hiperpolarización, es decir, disminución de la corriente oscura, así se reduce la liberación de neurotransmisor en la sinapsis (glutamato) y se genera una señal que dará como resultado la génesis de potenciales de acción en las células ganglionares.
La acción de la luz reduce la concentración de GMP cíclico, produciéndose el bloqueo en la entrada del sodio. Asimismo se produce un bloqueo en la entrada de calcio. El aumento en la concentración de sodio en el exterior activa lo que conocemos como BOMBA INTERCAMBIADORA DE SODIO-CALCIO, lo cual provoca la liberación de calcio al exterior desde el segmento externo. La reducción del calcio intracelular inhibe la granulatociclasa y activa la fosfodiesterasa, así se vuelven a abrir los canales para el sodio y calcio, recuperándose el foto-receptor tras la excitación lumínica, dejándolo listo para la llegada de un nuevo fotón.

Esquema de la Fototransducción en el Segmento Externo del Bastón
PROCESAMIENTO NEURONAL MEDIANTE LA CONVERGENCIA.
Al observar las conexiones de conos y bastones con otras neuronas de la retina, nos daremos cuenta que hay un nivel de convergencia diferente en cada uno de estos foto-receptores. La convergencia de los bastones es mayor que la de los conos. Se admite una media de 120 bastones convergiendo sobre una sola célula ganglionar, mientras que tan solo seis conos convergen sobre una ganglionar, esta reducción es incluso mayor en la fóvea, donde llegamos a tener conexiones unitarias, un cono una ganglionar, es decir ya no habría convergencia, sería una conexión directa en la fovea. Esta diferencia entre conos y bastones explica que la visión mediada por conos sea más precisa en la detección de detalles, mientras que los bastones tienen mejor sensibilidad al contraste y menos sensibilidad al detalle, ahora veremos esto con detalle.
Los bastones son más sensibles que los conos al contraste porque necesitan menos luz para generar una respuesta y, especialmente por el fenómeno de convergencia, que determina una sumación de las intensidades. Si tenemos dos células ganglionares que necesitan cada una de ellas 5 unidades de intensidad para activarse, y sobre la primera tenemos un conjunto de cinco bastones convergiendo sobre ella y sobre la segunda le llega la conexión de un solo cono, cuando lleguen estímulos de 1 unidad de intensidad, por ejemplo ocupando un área que activa 5 foto-receptores, los bastones se estimularan y cada uno de ellos enviará 1 unidad sobre la cl ganglionar de convergencia, sumando 5 unidades, el mínimo requerido para activarla, mientras que en el caso de los conos, al activarse, envían tan solo una señal de 1 unidad y, si el estímulo activa 5 conos, como cada uno conecta con una cl. ganglionar, siempre le llega a la cl. ganglionar 1 unidad, no hay efecto de sumación como en los bastones ya que no hay convergencia múltiple, por tanto ese estímulo sí excita la célula ganglionar de los bastones pero no las células ganglionares de los conos. Esto explica porque los bastones son más sensibles a la luz que los conos.
Cuando analizamos la capacidad de detectar detalles, vemos que el fenómeno de convergencia produce el efecto contrario. En los conos de la fóvea, gracias a que cada uno conexiona con una ganglionar, permite que la resolución de la información sea mayor, podemos analizar la imagen punto por punto mientras que los bastones, como convergen varios en una ganglionar, la información se diluye y la resolución baja. Esto determina el concepto de agudeza visual, o capacidad para percibir los detalles.
En la oscuridad, la agudeza visual disminuye porque los conos dejan de funcionar y la visión se debe sólo a la acción de los bastones, con menor capacidad de resolución que los conos, por ello vemos menos nítido.
PROCESAMIENTO NEURAL MEDIANTE EXCITACIÓN E INHIBICIÓN.
Todos los sistemas neuronales funcionan con redes neurales que forman circuitos, es decir, un conjunto de neuronas conectadas entre sí. Cuando estamos en la situación de la fóvea, donde cada cono tiene su ganglionar, la activación de un cono-ganglionar, no se ve afectada por la estimulación de otros conos contiguos, mientras que en la retina periférica, donde predominan los fenómenos de convergencia, la activación de un bastón o más bastones contiguos, sí determina variaciones en la ganglionar donde convergen así, cuantos más bastones se estimulen, mayor frecuencia de respuesta en la ganglionar, tal como se observa en la figura.

Los circuitos pueden complicarse, especialmente cuando aparecen neuronas excitadoras, como el ejemplo anterior, junto a neuronas inhibidoras, como en la figura siguiente, donde las ganglionares A y C bloquean a la ganglionar B. Si el estímulo cae sobre los receptores centrales, 3-4-5, se activa la ganglionar B y se da un efecto excitador pero, si el estímulo es mayor en superficie y se excitan los foto-receptores 2 y 6, se activa parcialmente las ganglionares A y C y producen un efecto inhibidor sobre la ganglionar B que reduce su activación. Si el tamaño del estímulo aumenta y activa a todos los receptores, se suman los 1 y 7 y se activa con mayor potencia las ganglionares A y C, con lo que el efecto inhibidor sobre la B, es más potente y la respuesta total es menor.
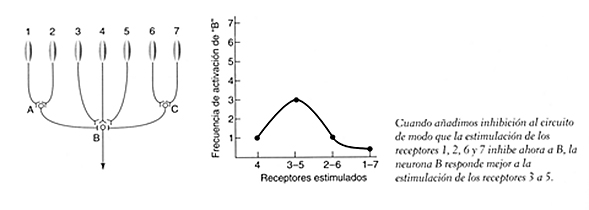
Recordemos que la retina ésta formada por diversos tipos de células y, la activación del receptor, cono o bastón, genera estímulos que llegan a las ganglionares a través de las células bipolares, horizontales y amacrinas, lo cual permite pensar que se pueden crear circuitos muy diversos, con vías de excitación o inhibición. Estas posibilidades permiten entender el concepto de campo receptor. En el ejemplo anterior, tendríamos un campo receptor correspondiente a una cel. ganglionar, en este caso la B, del tipo centro ON y periferia OF, también llamada on-of o centro-periferia, ya que si se estimula la zona central, se da una respuesta positiva de activación pero, si se actica la zona periférica, se inhibe la respuesta de la B y, si se activan todos los receptores de esta ganglionar B, se da una respuesta mínima, ya que la excitación de la zona central se ve inhibida por la zona periférica. Este tipo de respuesta, centro periferia, puede darse de la manera que acabamos de ver o, algo más complicada, las ganglionares A y C pueden no contactar directamente con la B, y hacerlo indirectamente mediante células de conexión, como las horizontales y amacrinas, que transmiten el efecto inhibidor, como el ejemplo anterior, pero nos permitirán entender mejor lo que conocemos como inhibición lateral, clave para la detección de contrastes.

La inhibición lateral fue demostrada en 1956 por Hatline, Wagner y Ratliff, utilizando el cangrejo de la especie Limulus y permitió demostrar como la estimulación de receptores adyacentes puede inhibir la respuesta de un receptor central, tal como se muestra en la figura.
PROCESAMIENTO NEURONAL Y PERCEPCIÓN.
El fenómeno de la inhibición lateral permite explicar fenómenos como el de la rejilla de Hermann, donde al mirar la rejilla en conjunto, vemos que en las intersecciones aparecen puntos grises, puntos que desaparecen si nos fijamos directamente en la intersección, demostrando que no son reales.

En la figura vemos que la ganglionar A, que está en medio de los dos pasillos, le llega inhibición de las 4 ganglionares que la rodean, mientras que la ganglionar B, que está en medio de un solo pasillo, le llega inhibición de las ganglionares que están en el pasillo, pero no de las ganglionares que están tapadas por los cuadrados negros, por ello la respuesta de la B es mayor que la de A y esto determina que en A se vea menos brillante y aparece como un punto gris en medio de la intersección de los cuatro cuadrados negros.

Otro de los efectos que se explican por la inhibición lateral son las Bandas de Mach. Tal como se muestra en la figura, si sobre una lámina iluminada colocamos algo que hace sombra, la línea divisoria de la zona en sombra respecto a la zona que sigue iluminada, no la percibimos como una línea bien definida sino que se aprecia una estrecha banda clara y, en la zona clara, junto a la línea divisoria, se ve una banda oscura, sin embargo, el análisis con fotómetro muestra que no hay tales bandas, registra una separación brusca y bien definida entre la zona más oscura y la más clara, lo que nosotros vemos, estas bandas de separación es un fenómeno subjetivo conocido como bandas de Mach y se deben al fenómeno de inhibición lateral.



Las bandas de Mach las podemos explicar con una representación en la que imaginamos 6 receptores, 3 estimulados por la zona clara y 3 por la zona oscura, de forma que cada receptor envía señales de inhibición lateral a los receptores contiguos. Si los receptores de la banda clara, supongamos que generan una respuesta de valor 100 y, los de la banda oscura, de valor 20. Si asumimos que cada célula envía una inhibición de una décima parte del valor de su respuesta, en las ganglionares de la banda clara, tendrán un valor de 10, mientras en las de la banda oscura, de 2. Si calculamos la respuesta final, resta de la activación menos la inhibición, en la ganglionar A, tendrá un valor de 100-10-10=80, en la B, igual, 80, pero en la C, 100-10-2= 88, en la D: 20-10-2= 8 y en E y F, 20-2-2= 16. En las ganglionares del borde, la inhibición es diferente, en la C, de la banda clara, hay menos inhibición, 88, respecto a los 80 de A y B, lo que hace que se vea más brillante y, en el otro lado, la ganglionar D, tiene mayor inhibición que las E y F, ya que recibe la inhibición de C, con valor de 10, lo cual hace que la respuesta final sea de 8, frente a los 16 de las otras, lo cual hace que se vea una franja más oscura.
El fenómeno de la inhibición lateral permite establecer un mecanismo fisiológico para hacer resaltar los bordes y también permite explicar los fenómenos de ilusiones ópticas como el contraste simultaneo de claridad, la cruz de Benary o la ilusión de White.
935513300 - info@areaoftalmologica.com
No hay comentarios:
Publicar un comentario